
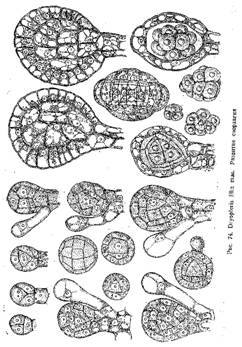
Теперь весь зачаток состоит из пяти клеток— одной пирамидальной, внутренней, и четырех наружных, окружающих ее. Внутренняя клетка является археспориальной клеткой, и в нее развивается содержимое спорангия — опоры и выстилающий слой. Наружные же дают начало стенке спорангия. Они делятся только радиальными перегородками, так что стенка спорангия все время остается однослойной. В археспориальной же клетке появляются перегородки, тангенгальные, параллельные ее граням. Отделившиеся таким образом четыре таблитчатые клетки дают начало тапетуму, внутренняя же является опорообразующей, из нее развиваются споры. Клетки тапетума снова делятся тангентальными перегородками, вследствие чего он становится двуслойным. В то же время делится и опорообразующая клетка, постепенно распадаясь на 16 клеток. Вскоре стенки клеток тапетума расплываются и протопласты их сливаются, образуя периплазмодий. Клетки же археспория обособляются друг от друга и округляются, превращаясь в материнские клетки опор, которые обычным путем дают начало спорам.
Евопорангиатный тип спорангиев встречается у наиболее древних и примитивных групп среди ныне живущих папоротникообразных. Он же характерен) и для ископаемых папоротникообразных. Это заставляет нас смотреть на евшорангий как на древний, примитивный тип спорангия, от которого уже произошел лептосггорангий, свойственный к тому же более молодым группам папоротников (Filices). Об этом говорит и то, что, как показывают сравнительные исследования, между евопорангиатный я лептоспорангиатным типами развития существует ряд постепенных переходов, связывающих их между собой. Вместе с тем эти переходы указывают на ход эволюции лептоспорангия.
На рис. 75 дана схема заложения опорангия у различных папоротников: рис. а относится к типичному лаптоспорангию (сем. Polypodiaeeae), а рис. g— к типичному евспорангию (Angiopteris). Между ними располагаются промежуточные типы, принадлежащие к различным родам. Понятно, эволюция лептоспорангия совершалась в направлении, обратном расположению рисунков. У Angiopteris археспорий закладывается глубоко под поверхностью и самая археспориальная клетка имеет форму куба: ограничивающие ее стенки пересекаются под прямым углом. У Todea (сем. Osmundaceae) наблюдаются два варианта развития опорангия. В одном случае ([) археспорий имеет в разрезе трапециевидную форму, благодаря тому, что антиклины, ограничивающие археспорий, наклонены под некоторым углом друг к другу. В другом случае (более мелкие спорангии) эти антиклины пересекаются, и археспорий принимает форму четырехгранной пирамиды, обращенной вершиной вниз, то есть форму, характерную для археспория лептоепорангия. И у Angiopteris и у Todea в образовании спорангия принимают участие и прилежащие к археспорию клетки. Развитие спорангия у Schiziaea (d), Alsophila(с) и Ceratopteris (Ь) представляет переход к типичному лептоспорангиатному типу (а). Археспорий окончательно принимает форму четырехгранной пирамиды и все меньшее число эпидермических клеток идет на развитие спорангиев. У Polypo-diaceae (а) весь спорангий развивается из одной эпидермической клетки.
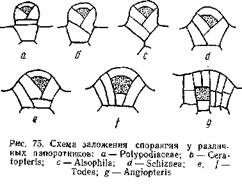
Таким образом, эволюция спорангия у папоротников шла в направлении редукции, которая и приведет к образованию лептоспорангия. Эволюция последнего, кроме того, выразилась в создании длинной и тонкой ножки, выработке кольца и в общем уменьшении количества производимых спорангием спор. В результате получился типичный лептоспорангий малой величины с длинной ножкой и однорядным кольцом. Число спор, образующихся в спорангии, невелико, но число спорангиев сильно возрастает, благодаря чему общее количество спор, производимых растением, не только не уменьшается, но и весьма увеличивается.
По характеру спор среди археголиальных мы различаем две труппы растений — равноепоровые и разноюлоровые (гомо- и гегероспоровые) формы. И те и другие встречаются во всех классах папоротникообразных; голосемянные—исключительно тетероспоровые. Это показывает, что гетероспория возникала в разных типах и классах самостоятельно я независимо. Естественно встает вопрос, какой тип спор мы должны считать первоначальным, исходным, иначе, развилась ли гетероспория из гомоспории или, наоборот, гетероспория дала начало гомоспории. Прямых указаний на то или иное решение вопроса мы не имеем, теоретически же возможно допустить и тот и другой ход эволюции. Однако, исходя из того факта, что гетероспория всегда связана с сильной редукцией гвметофита, в принимая во внимание то, что примитивный гаметофит должен был быть хорошо развитым образованием, правильнее будет предположить, что гомоопория является примитивным признаком и первоначально все опоры были гомоспорами, то есть при прорастании давали обоеполые заростки, несущие и антеридии и архегонии. В дальнейшей эволюции произошла дифферевцировка их на микроспоры и мегаспоры — мужские и женские опоры. Указание на такого рода дифференцирозку мы находим и среди мхов, согласно исследованиям сравнительно недавнего времени.
Статьи и публикации:
Понятие о коренных, или климаксовых растительных сообществах. Серии, стадии,
производные фитоценозы
В процессе сукцессий возникают биогеоценозы, в наибольшей степени соответствующие условиям среды, как климатическим, так и эдафическим, а также состоящие из «подогнанных» к совместному обитанию видов со свойственным этому ценозу фитоклима ...
Охрана труда и техника
безопасности на предприятиях, выпускающих ферментные препараты
Инженерные мероприятия являются наиболее важными. Они призваны максимально предотвратить выход и вынос ферментных препаратов в помещения и за пределы предприятия, т. е. исключить контакт с ними человека. Известно также, что в производстве ...
Предмет и структура естествознания. Понятие естествознания
Стремление человека к познанию окружающего мира выражается в различных формах, способах и направлениях исследовательской деятельности. Каждая из основных частей объективного мира — природа, общество и человек — изучается своими отдельными ...