
Можно говорить о множественной роли процессов катаболизма.
Во-первых, роль корректирующего фактора (рис. 1). Он обеспечивает, устранение биополимеров с «некорректной» структурой, вызванной тремя возможными причинами: ошибками в ходе образования этих соединений; большей или меньшей потерей их активности в результате последующей модификации; таким изменением окружающей среды, при котором эти биополимеры или липиды не могут в полной мере выполнять свои функции.
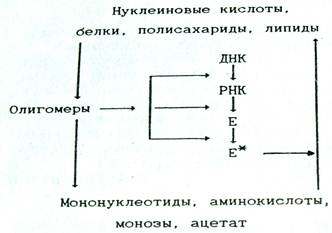
Рис. 1. Схема взаимосвязи катаболизма и анаболизма, отражающая субстратную и регуляторную функции катаболизма
Во-вторых, субстратная роль. Процессы катаболизма обеспечивают синтез биополимеров мономерными субстратами, а синтез липидов - ацетатом, что особенно важно в условиях недостаточного насыщения клеток этими веществами.
В-третьих, энергетическая роль. При дальнейшей деградации мономерных продуктов и ацетата и вовлечении их в процессы дыхания образуются АТФ и НАД(Ф)Н, обеспечивающие протекание анаболических реакций.
В-четвертых, сигнальная функция. Известно, что особенности структуры нуклеиновых кислот, белков и некоторых поли- и олигосахаридов (в том числе входящих в состав гликопротеидов и гликолипидов) используются в клетках для хранения, передачи и реализации информации.
В процессе деградации биополимеров происходит постепенное уменьшение заключенного в этих молекулах объема информации. Однако трудно себе представить, что в ходе эволюции не были использованы информационные свойства фрагментов молекул биополимеров, образующихся при деструкционном каскаде в результате действия эндо- и экзогидролаз. Исследования в этом направлении показали, что, во-первых, частичная деградация предобразованных молекул является широко распространенным способом формирования функциональных молекул нуклеиновых кислот и белков в реакциях соответственно посттранскрипционного процессинга РНК и посттрансляционной модификации белков.
Во-вторых, накапливается все больше информации о том, что некоторые олигонуклеотиды, олигопептиды и олигосахариды являющиеся продуктами катаболизма соответствующих полимеров обладав свойствами гормонов или активаторов и ингибиторов различных процессов метаболизма и в конечном итоге оказывают влияние на рост и морфогенез растений.
Можно считать, что сигнальные свойства олигомерных промежуточных продуктов катаболизма реализуются путем воздействия на транскрипцию, трансляцию или на предобразованные ферменты связанные с процессами анаболизма, а также на мембранные структуры (см. рис. 1)
Приведенные выше рассуждения о сигнальных свойствах олигомерных производных биополимеров справедливы и в отношении промежуточных продуктов деградации липидов, в состав которых входят полиеновые (ненасыщенные) жирные кислоты — линолевая, линоленовая, а у низших грибов и бактерий - арахидоновая и эйкоэапентаеновая, обладающие соответственно двумя, тремя, четырьмя и пятью двойными связями. Сигнальные свойства полиеновых жирных кислот определяются реакциями оксигенирования молекул по местам двойных связей и последующими реакциями, в результате осуществления которых появляется множество производных жирных кислот. Некоторые из этих продуктов оксигенирования являются физиологически активными веществами.
Деградация мембранных липидов может приводить к драматическим последствиям, в связи с влиянием на физико-химические свойства мембран и условия функционирования катализирующих разнообразные процессы мембранных белков-ферментов, транслокаторных систем, рецепторов и т.д.
Скорость деградации и окислительных реакций в мембранных липидах имеет большое значение для регуляторной и информационной роли мембран в клеточном метаболизме.
Некоторые из промежуточных продуктов метаболизма биополимеров и липидов в ходе эволюции животных и растительных организмов стали выполнять функции гормонов - физиологически активных веществ, вырабатываемых в одних органах, транспортируемых в другие и изменяющих в них интенсивность и направленность метаболизма и в конечном итоге влияющих на реализацию программ роста, морфогенеза, устойчивости и др.
Принципиальное отличие этих систем у растительных и животных организмов в том, что у первых отсутствуют полипептидные или олигопептидные гормоны, а в роли гормонов стали выступать менее сложные вещества, образующиеся, например, при модификации мономерных продуктов катаболизма белков (триптофан - ауксин, метионин – этилен) нуклеиновых кислот (пуриновые нуклеотиды - цитокинины) или липидов (жирные кислоты - жасмонат).
Большинство же физиологически активных олигомерных продуктов катаболизма биополимеров и оксигенированных производных полиеновых жирных кислот выполняет роль не гормонов, а внутриклеточных эндогенных регуляторов, не транспортируемых в другие органы и ткани. Они могут играть роль микровинта в корректировании метаболических процессов, в то время как грубая настройка осуществляется фитогормонами.
Статьи и публикации:
Отдел полезащитных насаждений
Отдел занимается разработками технологий выращивания лесонасаждений на базе широкой механизации с минимальными затратами ручного труда на богарных и орошаемых сельскохозяйственных землях, на оросительных системах, вдоль ирригационной сети ...
Представление о консорции: структура и функции. Роль биотических факторов в
формировании растительности
Сочетание разнородных организмов, тесно связанных друг с другом в их жизнедеятельности известной общностью их судьбы. Напр., древесная порода (ель, береза) со свойственными ей паразитами (лишайник), симбиозом (гриб), вредителями, переносч ...
Выход продукта
Организация «дробной» подпитки приводит к активизации биосинтетической деятельности микроорганизмов. На мелассной среде осуществление «дробной» подпитки позволяет увеличить выход до 60 г/л. Количество фосфора в среде должно быть в предела ...