
Тонкие механизмы регуляции уровня сАМР связаны с функционированием фосфотрансферазной системы транспорта Сахаров и будут рассмотрены в главе, посвященной регуляции процессов мембранного транспорта.
Необходимо отметить, что у ряда бактерий роль глюкозы в катаболитной репрессии могут выполнять другие источники энергии, которые в этом случае тормозят катаболизм глюкозы.
В индуцибельных оперонах возможны и другие типы положительной регуляции, независимой от сАМР. Например, в арабинозном опероне Escherichia coti арабиноза не просто инактивирует репрессор, но превращает его в положительный регулятор. Аналогичное явление обнаружено в случае оперонов галактозы и рамнозы.
В дивергентных регулонах транскрипция протекает в разных направлениях и может быть некоординированной, т.е. осуществляться с разной скоростью. При этом возможно считывание с разных цепей ДНК. Примером служит аргининовый оперон: в его части, включающей 4 гена из 9, транскрипция трех генов осуществляется в одном направлении, а транскрипция другого гена – в противоположном:
![]()
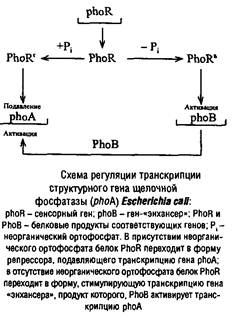
Еще одним примером положительной регуляции процесса транскрипции является регуляция с участием генов – «энхансеров». Ранее считали, что этот тип регуляции характерен только для эукариот. Но в последнее время формально сходные механизмы обнаружены и у прокариот.
Особенность генов – «энхансеров» в том, что они проявляют свою стимулирующую активность независимо от ориентации и расположения относительно активируемого гена: могут находиться перед геном, за ним и даже внутри него.
Продуктами генов – «энхансеров» являются белки с молекулярной массой 25–30 кДа, способные связываться с промоторной областью. Как правило, такая система двухкомпонентна и включает «сигнальный» белок, способный «чувствовать» изменение условий окружающей среды и стимулировать синтез другого белка – «активатора», который и запускает транскрипцию искомого белкового посредника.
Перечисленные механизмы регуляции транскрипции на стадии инициации достаточно быстро реагируют на изменение внешних условий, однако управляют работой одного или небольшого числа оперонов в каждый момент времени.
Наряду с ними существуют механизмы системной регуляции, связанные с изменением функционирования одновременно большого числа оперонов.
В клетках эукариот это достигается путем конформационных перестроек хроматина, процессинга и РНК, а также за счет управления трансляцией путем формирования так называемых информосом.
В клетках прокариот системная регуляция осуществляется путем модификации специфичности работы РНКП посредством изменения ее компонентного состава. Эти механизмы вступают в действие, когда нужно активировать одновременно большое число новых промоторов или сменить матрицу. Последний случай изучен наиболее подробно на примере фагов Escherichia coii и Bacillus subtilis.
В геноме бактериофагов присутствуют три типа генов: ранние, средние и поздние, классифицируемые на основании порядка их транскрипции в ходе развития фага. Ранние гены всегда транскрибируются РНКП клетки-хозяина, а в случае транскрипции средних и поздних генов возможно несколько вариантов:
Статьи и публикации:
Образование новых видов
С давних пор человека поражало многообразие органического мира. Как оно возникло? Учение о естественном отборе объяснило, как образуются новые виды в природе. Дарвин исходил из фактов, касающихся домашних пород. Первоначально породы домаш ...
Терминальная
дезоксинуклеотидил-трансфераза
а. Полимеризация без матрицы
Терминальная дезоксинуклеотидилтрансфераза является в определенном смысле полимеразой, поскольку она катализирует синтез полидезоксирибо-нуклеотидов из дезоксирибонуклеозидтрифосфатов с высвобождением неорган ...
Питательные среды и условия
культивирования
При конструировании оптимальных питательных сред для каждого продуцента изучаются его физиология, потребность в источниках N, С и других соединениях. Содержание сухого вещества в питательной среде в зависимости от продуцента может изменят ...