
Для проведения адаптации был выбран ступенчатый режим увеличения концентрации 2H2О в ростовых средах в присутствии 0.5-1% метанола/[U-2H]метанола , так как мы предположили, что постепенное привыкание организма к дейтерию будет оказывать благоприятный эффект на параметры роста и общее самочувствие культуры (табл. 1). При этом штамм M. flagellatum обнаружил повышенную чувствительность к 2H2O: ингибирование скорости роста бактерий наблюдалось при концентрации 2H2О в среде 74.5%, в то время как [U-2H]метанол не оказывал существенного влияния на скорость роста культуры. Так, на среде, содержащей 74.5% 2H2О выход микробной биомассы составил 29%, что в 3.4 раза ниже, чем в контрольных экспериментах, когда использовали обычную воду и метанол Какой мфу выбрать для офиса лучшие мфу для офиса.
(табл. 1, опыт 1), в то время как выход микробной биомассы на водной среде с 1% [U-2H]метанолом был снижен всего лишь в 1.2 раза. В связи с тем, что роста бактерий на более высокой концентрации 2H2O достичь не удалось в дальнейших экспериментах использовали биомассу M. flagellatum, полученную со среды, содержащей 74.5% 2H2О.
Адаптация факультативных метилотрофных бактерий B. methylicum.
Попытки адаптировать штамм B. methylicum к росту на максимально дейтерированной среде при сохранении способности к биосинтезу фенилаланина привели к положительному результату. К данному штамму метилотрофных бактерий был применён специальный подход по адаптации, который заключался в серии из нескольких адаптационных пассажей исходной культуры на твёрдых агаризованных средах с 2% [U-2H]метанолом при ступенчатом увеличении концентрации тяжёлой воды в них (от 0 до 98% 2H2О), как показано в табл. 2. При этом последовательно отбирали отдельные колонии, выросшие на средах, содержащих дейтерий. Затем их пересевали на среды с большей степенью дейтерированности, включая среду с 98% 2H2О (степень выживаемости бактерий на конечной максимально дейтерированной среде составила не более 50%). За ходом адаптации следили по изменениям продолжительности лаг-фазы, времени клеточной генерации и выходов микробной биомассы, а также по максимальному уровню накопления фенилаланина в культуральной жидкости (рис. 1).
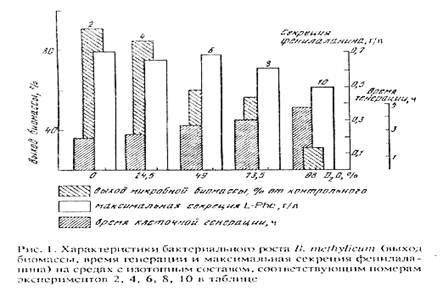
В отсутствии дейтериймеченных субстратов продолжительность лаг-фазы не превышала 20 ч, в то время как с увеличением концентрации 2H2О в ростовых средах до 98% продолжительность лаг-фазы увеличивалась до 60 часов (таблица 2, опыт 10). Отмечено, что длительность времени клеточной генерации с увеличением концентрации 2H2O в ростовых средах постепенно увеличивается, достигая 4,9 часов на среде с 98% 2H2O и 2% [U-2H]метанолом (табл. 2, опыт 10). В отличие от тяжёлой воды, [U-2H]метанол не вызывал существенного ингибирования роста и не оказывал влияния на выходе микробной биомассы. Напротив, на максимально дейтерированнной среде выход микробной биомассы был снижен в 3.3 раза по сравнению с контролем. Важно то, что выход микробной биомассы и уровень накопления фенилаланина в культуральной жидкости при росте адаптированного к 2Н2О микроорганизма в максимально дейтерированной среде изменяются по сравнению с контрольными условиями на 13 и 5%, т. е. незначительно (табл. 2, опыт 10’).
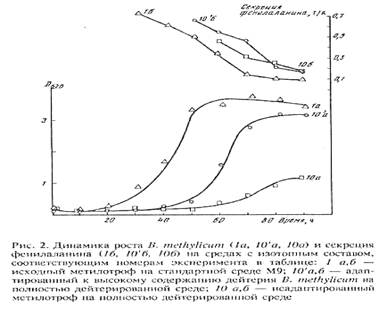
Адаптированные к 2Н2О клетки сохранили способность синтезировать и экзогенно продуцировать фенилаланин в ростовую среду. Причём общей особенностью биосинтеза фенилаланина в Н2О/2H2O-средах было увеличение его продукции на раней фазе экспоненциального роста B. methylicum, когда выход микробной биомассы был незначителен (рис. 2). Во всех экспериментах наблюдалось ингибирование биосинтеза фенилаланина на поздней фазе экспоненциального роста и снижение его концентрации в ростовых средах. Согласно данным по микроскопическому исследованию растущей популяции микроорганизмов, подобный характер динамики секреции фенилаланина не коррелировал с качественными изменениями ростовых характеристик культуры на различных стадиях роста, что служило подтверждением морфологической однородности микробной популяции. Скорее всего, накопленный в процессе роста фенилаланин ингибировал ферменты собственного пути биосинтеза. Кроме того, не исключена возможность, что при ферментации без рН-статирования может происходить как обратное превращение секретируемого фенилаланина в интермедиаторные соединения его биосинтеза, так и ассимиляция фенилаланина клеткой для обеспечения собственных метаболических потребностей, что отмечено в других работах [27, 28]. Данные по исследованию культуральной жидкости методом ТСХ показали, что кроме фенилаланина штамм B. methylicum синтезирует и накапливает в культуральной жидкости (на уровне 5-6 ммоль) другие аминокислоты (аланин, валин, лейцин, изолейцин), присутствие которых также подтверждалось масс-спектрометрическим анализом метиловых эфиров их DNS-производных.
Статьи и публикации:
Дайте общую характеристику кометам. Назовите
наиболее известные из них. Период обращения Сатурна вокруг Солнца равен 29,46
земного года, а Марса – 1,88 земного года. На каком расстоянии от Солнца
н
Кометы – это наиболее необычные по своему виду небесные объекты, которые иногда доступны для наблюдений невооруженным глазом. Они привлекали внимание человека с глубокой древности. Вместе с астероидами и метеорными телами их относят к мал ...
Классификация межклеточных контактов
1.Замыкающие межклеточные контакты.
а) простой или рыхлый контакт;
б) плотный замыкающий контакт.
2.Адгезионные межклеточные контакты.
а) точечные контакты;
б) адгезионные пояски;
в) адгезионные соединения между клеткой и внутриклет ...
Место проведения
исследования
Исследования по анализу пищевой ценности переработанных грибов проводили на Каменском консервном заводе - в производственной лаборатории. Производственная лаборатория является самостоятельным структурным подразделением предприятия и дейст ...