
Среди всех производных моносахаров, безусловно, на первое место следует поставить гликозиды. Гликозиды представляют собой моносахариды, в молекулах которых полуацетальный гидроксил замещен на какую-либо другую функциональную группу.
Из схемы 6.1 очевидно, что два атома кислорода при одном углероде как пиранозной, так и фуранозной форм моносахарида - это уже достаточное основание для создания существенного положительного заряда на этом атоме. Вследствие этого, для молекулы открываются две возможности реагирования: во-первых, облегчается диссоциация по связи С-ОН с образованием соответствующего карбкатиона, катионный центр которого стабилизирован взаимодействием с п-уровнем эндоциклического атома кислорода; во-вторых, достаточно большой положительный заряд на углероде создает благоприятную предпосылку для непосредственной атаки его нуклеофильным реагентом. Другими словами, ожидается, что полуацетальный гидроксил пираноз и фураноз будет легко замещаться нуклеофильно как по механизму SN1; так и по механизму SN2.
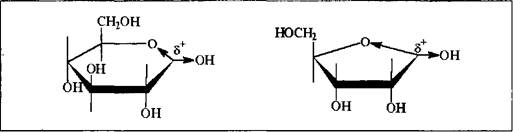
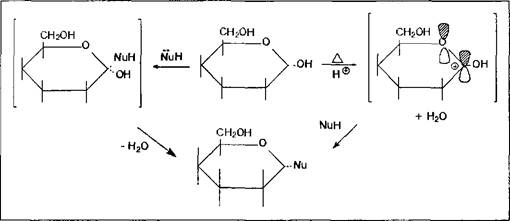
В соответствии с общими закономерностями реакций нуклеофильного замещения, в случае реализации мономолекулярного механизма ожидается образование продукта реакции в виде смеси диастереомеров, так как карбкатионный фрагмент переходного состояния молекулы плоский; если же процесс пойдет по синхронному бимолекулярному пути - пространственная структура продукта будет зависеть от конфигурации исходного моносахарида: из б-формы образуется в-гликозид, из в-формы - б-гликозид, так как атака нуклеофила осуществляется в тыл связи С-0 и завершается обращением конфигурации реакционного центра. Поскольку моносахарид всегда будет существовать в растворе в виде таутомерной смеси а - и в-форм, то независимо от механизма реакции мы, как правило, получим гликозид в виде изомерной смеси. Только лишь их соотношение будет варьироваться в зависимости от условий реакций и природы реагента. Но это все верно тогда, когда реакция проводится в колбе, т.е. in vitro.
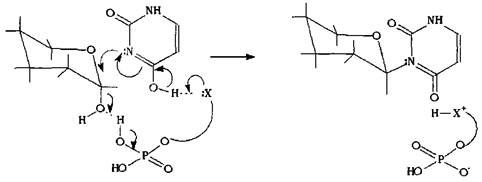
Внутри живой клетки такой процесс катализируется ферментами, которые, как правило, работают стереоспецифично: это значит, что они будут избирательно вовлекать в реакцию какой-либо один из диастереомеров, а также осуществлять реакцию по какому-либо одному механизму, что в итоге приведет к продукту одной изомерной формы. Схематично такую реакцию можно представить, моделируя реагент и фермент в виде единой молекулы, на одном конце которой находится остаток фосфорной кислоты, катализирующий отщепление гидроксильной группы, на другом конце - нуклеофильный остаток, атакующий электрофильный атом углерода. Синхронное воздействие каталитического и нуклеофильного фрагментов на электрофильный центр а-глюкопиранозы приводит к соответствующему в-гликозиду.
Вполне возможно, что б-изомер большинства моносахаров более реакционно способен хотя бы по причине стерической доступности его реакционного центра. Тогда следует ожидать, что большинство природных гликозидов будут относится к ряду в-изомеров, так как их образование ферментативно катализируемо. Трудно проанализировать
все природные гликозиды на предмет принадлежности их к а - или в-ряду в виду их многочисленности. Но все-таки создается впечатление, что в-диастереомеры все же более многочисленны. Так как б-изомерные формы часто также образуются исключительно стереоспецифично, следует предположить, что комплексирование каталитического сайта фермента и молекулы моносахарида чувствительно к исходной пространственной форме пиранозы или фуранозы. Это возможно только при условии, что образование одних и тех же гликозидов различной конфигурации должно катализироваться различными ферментативными системами.
Для всех гликозидов характерно отсутствие таутомерных превращений в растворе, т.е. переход их в ациклическую форму невозможен, так как для этого процесса требуется подвижный атом водорода полуацетального гидроксила для преобразования последней в карбонильную группу. Второе общее свойство гликозидов - это достаточно легкая способность их к гидролизу в кислой среде, фактически являющемуся процессом, обратным реакции их образования.
Классификация гликозидов. Классифицируются гликозиды достаточно однозначно - согласно типу нуклеофила, заместившего полуацетальный гидроксил. Такой нуклеофил также называют агликоном. Если агликонами являются спирты или фенолы, то образующиеся гликозиды называют 0-гликозидами; серусодержащие нуклеофилы ведут к образованию S-гликозидов; если нуклеофилом являлось соединение, генерирующее в ходе реакции карбанион, то полученный гликозид имеет структуру С-гликозида; и наконец, очень распространены в природе различные варианты образования Н-гликозидов в ходе замещения полуацетального гидроксила моносахаров азотистыми основаниями разного типа.
Статьи и публикации:
Ведение
экскурсии
В плодовом саду обитает ряд насекомых, которых можно с успехом использовать в качестве экскурсионного материала. Конечно, на юге насекомых, развивающихся за счет плодовых деревьев, значительна больше, но и в нашем северном плодовом саду м ...
Годичные изменения численности мелких млекопитающих
Накопленный в настоящее время материал по разным группам животных организмов показывает, что численность естественных популяций не остаётся постоянной даже при выходе на плато логистической кривой. Более того, наряду с незакономерными и н ...
Расчет количества корма ОСТ – 4 для молоди осетровых
Личинка m = 60 мг.
35% от массы тела необходимо корма в сутки.
Общая биомасса 126 кг.
Затрачивается 44,1 кг. корма в сутки.
Т.к. изначально личинку подкармливают науплиями артемии салина количество комбикорма сокращают на 50% и его ма ...