
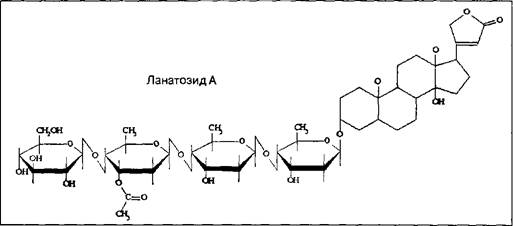
Примером таких гликозидов может служить ланатозид А, найденный в одном из видов наперстянки.
Другая интересная группа растительных О-гликозидов - это цианогенные гликозиды, агликоновый компонент которых образован из а-циангидринов. Их особенностью является способность выделять синильную кислоту при ферментативном гидролизе, которому они подвергаются чрезвычайно легко: появление запаха "горького миндаля" при использовании семян некоторых растений указывает на присутствие в них таких гликозидов и на начало реакции их распада.
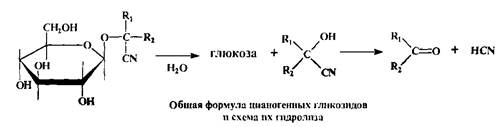
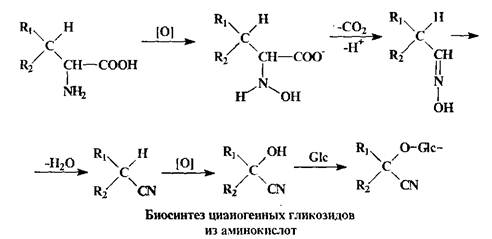
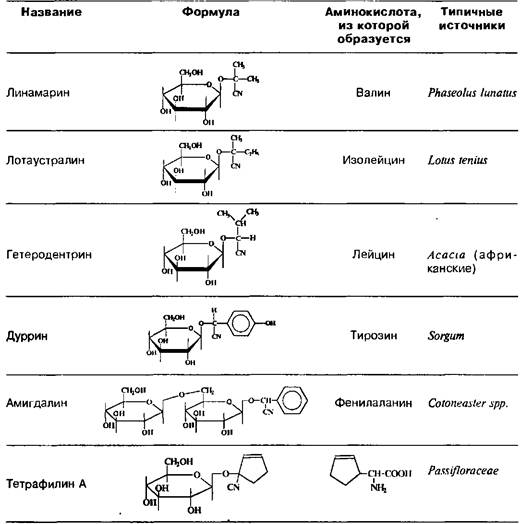
Циангидрины цианогенных гликозидов образуются из соответствующих аминокислот; в качестве сахарного остатка почти всегда фигурирует D-глюкоза, гликозидная связь которой имеет в-конфигурацию.
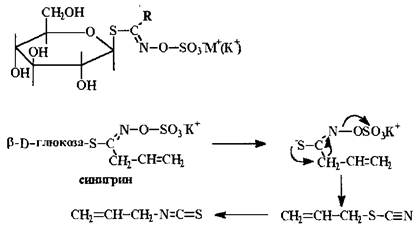
S-гликозиды. S-гликозиды весьма немногочисленны, наиболее хорошо изучена группа глюкозинолетов. Эти лечебные соединения легко расщепляются при действии ферментов, выделяя соответствующие изотиоцианаты и тиоцианаты, являющиеся причиной отравления при использовании растений, содержащих эти гликозиды.
Они содержатся во многих видах крестоцветных, максимум накопления тиогликозидов у большинства из них - в недозрелых семенах.
Н-гликозиды. О степени важности для живых систем Н-гликозидов говорить не приходится. Нуклеиновые кислоты, многие коферменты, макроэрги - вот те классы природных веществ, в основе которых лежит Н-гликозидный фрагмент, играя при этом далеко не второстепенную роль. Можно утверждать, что нет живой клетки без Н-гликозидов. Но при всем при этом, в структурном плане они представляют собой довольно локальный класс природных соединений, так как он ограничен сравнительно небольшим набором азотистых агликонов, а еще в большей степени - набором моносахаров, участвующих в их формировании. Поскольку основная масса Н-гликозидов представлена фрагментами, составляющими базис нуклеиновых кислот, и именно эти гликозиды выделены на первичных этапах исследования состава и строения ДНК и РНК, то из них и образовали отдельную группу под названием нуклеозиды.
Нуклеозиды образованы всего двумя моносахаридами - D-рибозой и D-2-дезокси-рибозой в своих фуранозных формах и небольшой серией азотистых гетероциклов с в-конфигурацией гликозидного фрагмента.
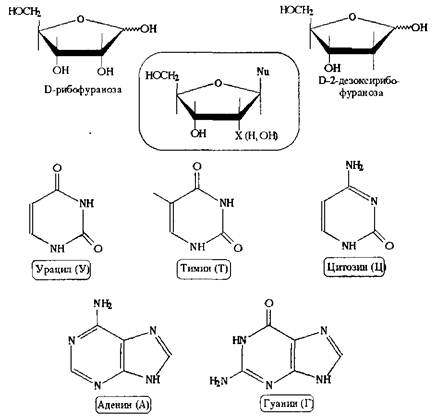
Азотистые агликоны нуклеозидов представлены двумя группами гетероциклов: производными пиримидина и производными пурина. Пиримидин и пурин функционализированы - НЗ2 и ОН-группами, но последний функционал претерпевает таутомерное превращение из гидроксиформы в соответствующую карбонильную функцию - это равновесие в нейтральной среде сдвинуто в сторону пиридоновых форм, тогда как соли имеют гидроксипиридиновую структуру.
Аминопиридиновый фрагмент этих нуклеиновых оснований также способен к таутомерному превращению, равновесие обычно сдвинуто в сторону аминной формы.
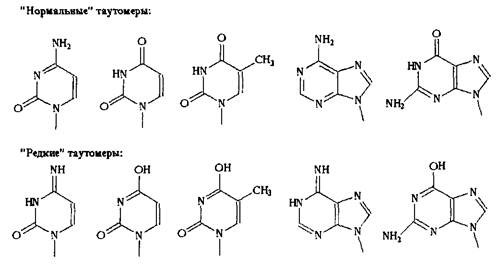
Таким образом, нуклеозиды можно разделить на "нормальные" - те, которые содержат в гетероцикле карбонильные и аминные функции, и на "редкие" - те, которые содержат в гетероцикле гидроксильные и иминные функции.
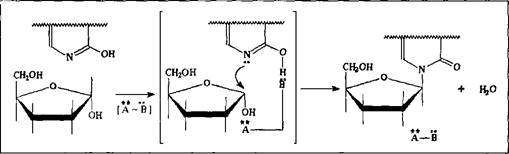
Механизм образования Н-гликозидов этого ряда может быть объяснен, основываясь на следующих предпосылках: как правило, моносахара в водных растворах предпочтительно существуют в б-форме; во-вторых, все нуклеозиды при этом имеют в-конфигурацию; и, наконец, поскольку образование Н-гликозида - это процесс нуклеофильного замещения, из двух таутомерных форм всех нуклеиновых оснований нуклеофильными свойствами обладают их лактимные формы. Учитывая все это, можно предположить, что реакция идет по механизму SN2, при котором электрофильный центр С атакуется атомом азота в пиридиновом валентном состоянии. А если учесть еще и то, что процесс должен реализоваться в очень мягких условиях живой клетки, можно предположить двойной катализ этой реакции - кислотный по полуацетальному гидроксилу и основной по фенольному гидроксилу. В принципе, он может быть реализован одной молекулой фермента, имеющей и кислотную, и основную функциональную группу.
Аналогичным образом могут быть образованы Н-гликозиды - с участием аминокислот и других биогенных аминов. Так как аминокислоты в нейтральных условиях существуют преимущественно в цвиттер-ионной форме, то катализатор-фермент двойственной природы, связывая кислотную функцию, активирует тем самым нуклеофильный реагент - а его кислотная функция будет синхронно активировать электрофильный центр.
Статьи и публикации:
Человеческая деятельность и принцип экологической
эквивалентности
Человек, воздействуя на экосистемы и отторгая часть вещества и энергии в производственный цикл, нарушает биотические круговороты, что неминуемо сказывается на состоянии окружающей среды. Как правило, она становится неблагоприятной для жиз ...
Характеристика семейства астровых (сложноцветные). Четыре типа цветков. Наиболее
распространенные растения из этого семейства: кормовые, сорные, ядовитые,
декоративные, лекарственные
А́стровые
(лат. Asteráceae), или Сложноцве́тные
(лат. Compósitae) — одно из самых больших семейств двудольных растений; включает около 25 тысяч видов (относящихся к 900—1000 родам), распространённых по всему земн ...
Концентрирование ферментных
растворов методом вакуум-выпаривания
Экстракты из поверхностных культур микроорганизмов и фильтраты глубинной культуры являются нестабильными при хранении. Для получения готовых форм технических препаратов (П2х и Г2х) их необходимо сконцентрировать. Чаще всего для этих целей ...