
морщинистыми зелеными семенами, то они всегда гомозиготны и представлены одним генотипом aabb. Таким образом, число различных генотипических комбинаций у второго поколения гибридов (F2) оказывается равным девяти.
Рассмотренные количественные отношения между числом различных фенотипов и генотипов в F2 при дигибридном скрещивании справедливы для аллелей с
полным доминированием. Ясно, что в случаях промежуточного характера наследования число фенотипически различных форм будет больше. Если по 6 обоим признакам наследование промежуточное, то количество фенотически различных групп равняется числу генотипически различных групп.
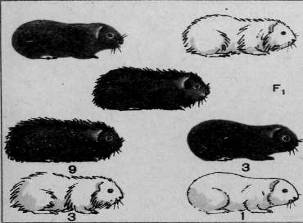
Рис. 1. Скрещивание и ход расщепления в двух поколениях двух пород морских свинок, различающихся по двум аллелям – окраске и форме шерсти (дигибридное скрещивание).
Ход дигибридного скрещивания можно показать и на примере животных. На рисунке 1 изображено дигибридное скрещивание двух пород морских свинок: черных гладких с белыми мохнатыми. В данном случае черная окраска доминирует над белой, мохнатая шерсть — над гладкой. Из рисунка без дальнейших пояснений ясен ход скрещивания, который совершенно аналогичен рассмотренному дигибридному скрещиванию горохов.
Второй закон Менделя.
Сопоставим результаты дигибридного и моногибридного скрещиваний. Если учитывать результаты расщепления по каждой аллели в отдельности, то легко видеть, что соотношение, характерное для моногибридного скрещивания, сохраняется. При рассмотренном выше дигибридном расщеплении у горохов отношение числа желтых семян (А) к зеленым (а) равняется 12:4 (3:1). То же касается и отношения гладких семян (В) к морщинистым (b). Таким образом, дигибридное расщепление представляет собой, по существу, два независимо идущих моногибридных, которые как бы накладываются друг на друга. Это может быть выражено алгебраически как квадрат двучлена (3+1)2 = 32 + 2хЗ+12, или, что то же самое, 9+3 + 3+1. Мы подошли, таким образом, к формулировке второго очень важного закона, установленного Менделем, который можно назвать законом независимого распределения генов. Он гласит, что расщепление по каждой паре признаков (по каждой аллели) идет независимо от других пар признаков (относящихся к другим аллелям).
Цитологические основы дигибридного расщепления.
Как связать закономерности дигибридного расщепления с теми процессами, которые совершаются в половых клетках при их созревании и оплодотворении? Эти отношения поясняются на схеме (рис. 106). Диплоидный набор хромосом представлен здесь двумя гомологичными парами. В парных хромосомах расположены аллельные гены. В палочковидных хромосомах гены А (красные) и а (синие), в сферических хромосомах гены В (красные) и b (синие). В результате мейоза из каждой гомологичной пары хромосом в гаметах остается по одной (см. схему). В результате оплодотворения в двойной гетерозиготе АаВb в каждой паре хромосом будут разные гены одной аллели (на схеме красная и синяя). При редукционном делении у гибрида первого поколения (F1) в равном количестве образуется четыре сорта гамет. Это зависит от того, что при мейозе во время конъюгации хромосом, предшествующей их расхождению, взаимное расположение хромосом разных гомологичных пар носит случайный характер. Если, например, к одному полюсу отходит «синяя» палочковидная хромосома, то из другой пары с одинаковой долей вероятности может отойти или тоже «синяя», или же «красная». В результате оплодотворения и развития второго поколения гибридов (F2) одинаково вероятно образование 16 категорий зигот. На схеме все возможные комбинации хромосом в зиготах врисованы в квадрате.
Статьи и публикации:
Охрана труда и техника
безопасности на предприятиях, выпускающих ферментные препараты
Инженерные мероприятия являются наиболее важными. Они призваны максимально предотвратить выход и вынос ферментных препаратов в помещения и за пределы предприятия, т. е. исключить контакт с ними человека. Известно также, что в производстве ...
Получение наночастиц платины
Маленькие частицы платины, представляющие интерес для каталитических процессов, можно получить путем восстановления водорастворимого комплекса в микроэмульсиях «вода в масле». Гексахлороплатиновая кислота растворяется в микрокаплях воды м ...
Природа
Природа Самарской области, с одной стороны, типична для средней полосы России, с другой стороны, воистину уникальна. Особенность ее состоит в том, что на сравнительно небольшой территории можно найти элементы природных зон, характерных пр ...