
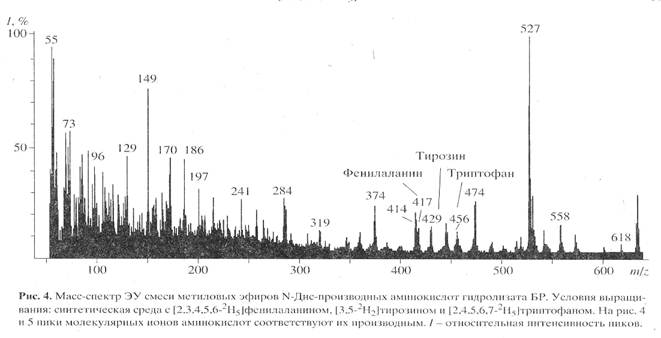
Для изучения уровня дейтерированности 2H-меченого БР использовали метод масс-спектрометрии электронного удара (чувствительность 10-8-10-10 моль анализируемого вещества [18]) после модификации смеси свободных аминокислот гидролизата БР в метиловые эфиры N-Днс-производных аминокислот. Чтобы получить воспроизводимый результат по уровню дейтерированности 2Н-меченого белка, сначала регистрировали полный скан масс-спектр электронного удара смеси метиловых эфиров N-Днс-производных 2Н-меченых аминокислот, по пикам молекулярных ионов которых (М)+ рассчитывали уровень дейтерированности молекулы. Затем проводили разделение метиловых эфиров N-Днс-производных ароматических аминокислот обращенно-фазовой ВЭЖХ и получали масс-спектры электронного удара для каждой индивидуальной аминокислоты. Полный масс-спектр электронного удара смеси метиловых эфиров N-Днс-производных аминокислот, показанный на рис. 4 (сканирование при m/z 50-640, базовый пик m/z 527, 100%), отличался непрерывностью, пики в интервале m/z от 50 до 400 на шкале массовых чисел представлены фрагментами метастабильных ионов, низкомолекулярных примесей, а также продуктами химической модификации аминокислот. Анализируемые 2Н-меченые ароматические аминокислоты, занимающие шкалу массовых чисел m/z от 415 до 456 представлены смесями молекул с различным количеством включенных атомов дейтерия, поэтому молекулярные ионы (М)+ полиморфно расщеплялись на отдельные кластеры со статистическим набором значений m/z зависимости от количества водородных атомов в молекуле. Учитывая эффект изотопного полиморфизма, подсчет уровня дейтерированности молекул аминокислот проводили по наиболее распространенному пику молекулярного иона (М)+ в каждом кластере с математически усредненной величиной (М)+ (рис. 4) - для фенилаланина пик молекулярного иона определялся (М)+ при m/z 417, 14% (вместо (М)+ при m/z 412, 20% для немеченого производного (пики немеченых аминокислот не показаны)), тирозина - (М)+ при m/z 429, 15% (вместо (М)+
при m/z 428, 13%), триптофана - (М)+ при m/z 456, 11% (вместо (М)+ при m/z 451, 17%). Уровень дейтерированности, соответствующий увеличению молекулярной массы составил для тирозина два, фенилаланина и триптофана - пять атомов дейтерия. Полученные данные по уровню дейтерированности фенилаланина, тирозина и триптофана позволяют сделать вывод о высокой селективности включения 2H-меченых ароматических аминокислот в молекулу БР: дейтерий детектировался во всех остатках ароматических аминокислот (таблица). Обсуждая полученные результаты, необходимо подчеркнуть, что присутствие в масс-спектре пиков (M)+ протонированных и полудейтерированных аналогов фенилаланина с (M)+ при m/z 413-418, тирозина с (M)+ при m/z 428-430 и триптофана с (M)+ 453-457 с различными вкладами в уровни дейтерированности молекул, свидетельстствует о сохранении небольшой доли минорных путей биосинтеза de novo, приводящим к разбавлению дейтериевой метки и, по-видимому, определяется самими условиями биосинтеза 2Н-меченного БР (таблица).
Согласно данным масс-спектрометрического анализа, пики молекулярных ионов (М)+
метиловых эфиров N-Днс-производных ароматических аминокислот обладали очень низкой интенсивностью и полиморфно расщеплялись, поэтому области их молекулярного обогащения были сильно уширены. Кроме этого, масс-спектры компонентов смеси аддитивны, поэтому смеси можно анализировать, только если имеются спектры различных компонентов, записанные в тех же условиях [8]. Проводимые вычисления предусматривают решение системы из n уравнений с n неизвестными для смеси из n компонентов. Для компонентов, концентрация которых превышает 10 мол.%, правильность и воспроизводимость результатов анализа составляет +0.5 мол.% (при доверительной вероятности 90%). Поэтому для получения воспроизводимого результата необходимо хроматографически выделять индивидуальные производные 2Н-меченых аминокислот из белкового гидролизата. Для решения поставленной задачи использовали метод обращенно-фазовой ВЭЖХ на октадецилсилановом селикагеле силасорб С18, эффективность которого подтверждалась разделением смеси метиловых эфиров N-Днс-производных 2Н-меченых аминокислот из других микробных объектов, как метилотрофные бактерии и микроводоросли [19]. Метод удалось адаптировать к условиям хроматографического разделения смеси метиловых эфиров N-Днс-производных аминокислот гидролизата БР, заключающийся в оптимизации соотношения элюентов, форме градиента и скорости элюции с колонки. Наилучшее разделение достигалось при градиентной элюции метиловых эфиров N-Dns-производных аминокислот смесью растворителей ацетонитрил : трифторуксусная кислота = 100 : 0.1 - 0.5, об.%. При этом удалось разделить триптофан и трудно разрешимую пару фенилаланин/тирозин. Степени хроматографической чистоты выделенных метиловых эфиров N-Днс-[2, 3, 4, 5, 6-2H5]фенилаланина, N-Днс-[3, 5-2H2]тирозина и N-Днс-[2, 4, 5, 6, 7-2H5]триптофана составили 89, 91 и 90% при выходах 78-85%. Полученный результат подтвердил рис. 4, б на котором приведен масс-спектр электронного удара метилового эфира N-Днс-[2, 3, 4, 5, 6-2H5]фенилаланина, выделенного обращенно-фазовой ВЭЖХ (сканирование при m/z 70-600, базовый пик m/z 170, 100%) (масс-спектр приведен относительно немеченого метилового эфира N-Днс-фенилаланина (а), сканирование при m/z 150-700, базовый пик m/z 250, 100%). Доказательством включения дейтерия в молекулу фенилаланина является пик тяжелого молекулярного иона метилового эфира N-Днс-фенилаланина ((М)+ при m/z 417, 59% вместо (М)+ при m/z 412, 44% для немеченого производного фенилаланина) и дополнительный пик бензильного фрагмента фенилаланина С7Н7+ при m/z 96, 61% (вместо m/z 91, 55% в контроле (не показан)) (рис. 5, б). Пики второстепенных фрагментов различной интенсивности со значениями m/z 249, 234 и 170 принадлежат к продуктам вторичного распада дансильного остатка до N-диметиламинонафталина, низкоинтенсивный пик (M - COOCH3)+ при m/z 358, 7% (m/z 353, 10%, контроль) является продуктом отщепления карбоксиметильной СООСН3-группы из метилового эфира N-Днс-фенилаланина, а пик (M + CH3)+ при m/z 430, 15% (m/z 426, 8%, контроль) - продуктом дополнительного метилирования по a-аминогруппе фенилаланина (рис. 5, б). Согласно данным масс-спектра, разница между молекулярной массой легкого и тяжелого пиков [M]+ метилового эфира N-Днс-фенилаланина составляет пять единиц, что совпадает с полученными ранее данными по уровню дейтерированности исходного [2, 3, 4, 5, 6-2H5]фенилаланина, добавляемого в среду выращивания (масс-спектрометрические данные по уровням дейтерированности [2, 3, 4, 5, 6-2H5]фенилаланина, [3, 5-2H2]тирозина и [2, 4, 5, 6, 7-2H5]триптофана подтверждены спектроскопией 1Н ЯМР и находятся в корреляции).
Статьи и публикации:
Предмет и содержание анатомии и физиологии. Значение этих
наук в психолого-педагогической подготовке будущего учителя
Анатомия человека(от греч. aná — вверх и tomé — режу) — раздел биологии, изучающий морфологию человеческого организма, его систем и органов. Предметом изучения анатомии человека являются форма и строение, происхождение и раз ...
Структура капель микроэмульсий, содержащих белки
Большинство ферментативных реакций в микроэмульсиях проведено в системах типа «вода в масле», в которых фермент заключен в доменах воды. Систему можно представить следующей картиной: капельки воды диспергированы в масле, причем некоторые ...
Органы, системы и аппараты органов. Целостность
организма
Целостность организма человека, взаимообусловленность формы и функций его органов и систем подтверждаются при изучении взаимосвязи общих и местных нарушений в организме, возникающих при аномалиях зубочелюстной системы. В многочисленных ис ...